Haplogruppe R (Y-DNA)
| Haplogruppe des Y-Chromosoms | |
|---|---|
| Name | R |
| Möglicher Ursprungsort | Nordwestasian, Caucasus oder Südasien |
| Mutationen |
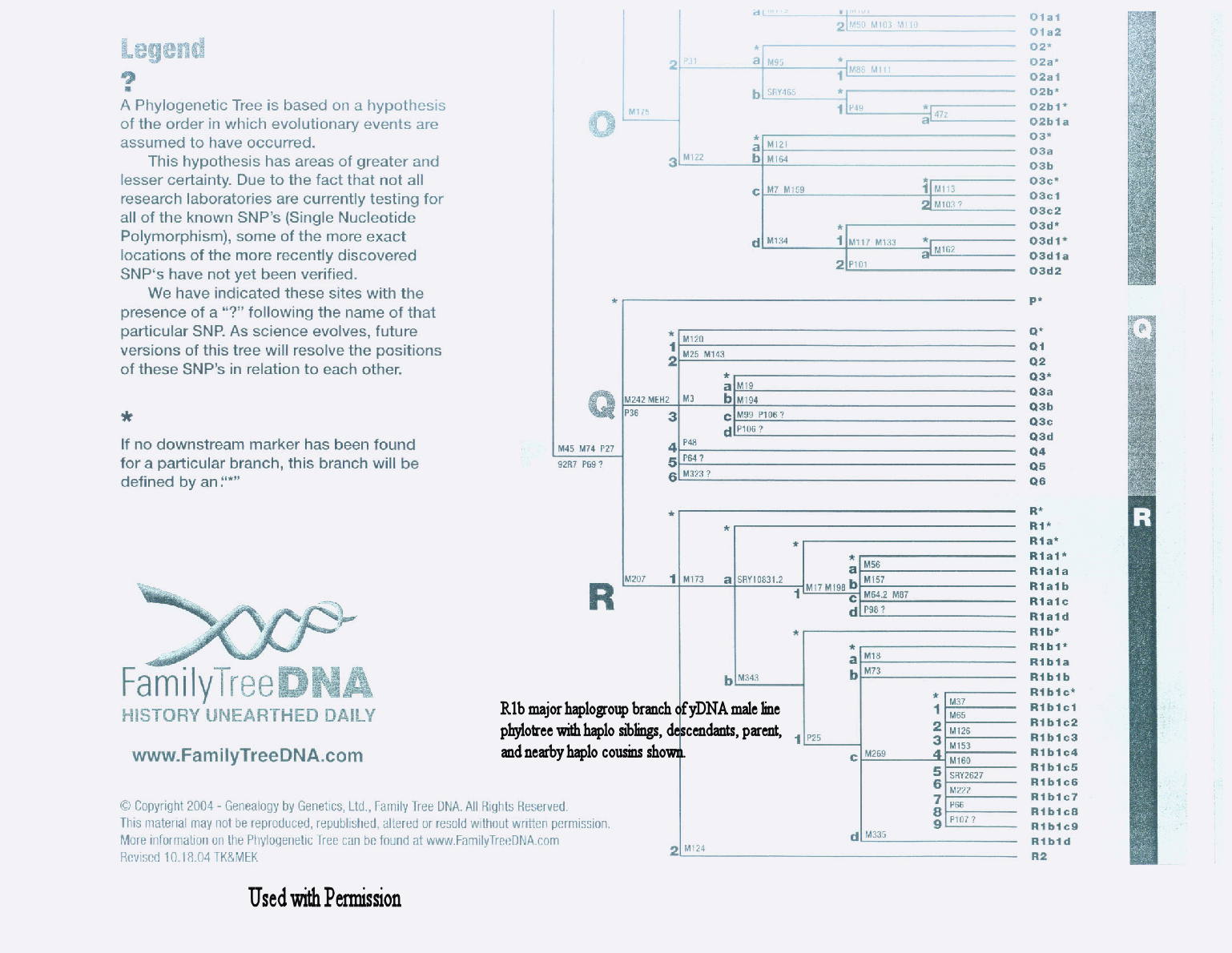
R = M207 (UTY2), P224, P227, P229, P232, P280, P285, S4, S8, S9 and V45 |
Die Haplogruppe R des Y-Chromosoms ist eine Haplogruppe des menschlichen Y-Chromosoms und eine Untergruppe der Haplogruppe P. Die Mutation M207 trägt zu ihrer Unterscheidung bei.
Ursprung
[Bearbeiten | Quelltext bearbeiten]Es wird angenommen, dass die Haplogruppe R in Nordwestasien vor 30.000 bis 35.000 Jahren erstmals auftrat. Sie stammt von der Haplogruppe P ab, welche ursprünglich von Ost-Asiatischen Jägern und Sammlern verbreitet wurde.[1] Sie ist die häufigste Haplogruppe in Europa und ebenso in Süd-, Zentralasien und im Kaukasus verbreitet. Seltenere Untergruppen finden sich in Australien, in Sibirien, bei amerikanischen Ureinwohnern, in Ägypten und Kamerun.
Verbreitung der Untergruppen
[Bearbeiten | Quelltext bearbeiten]Die Mehrheit der Träger von Haplogruppe R gehören der Haplogruppe R1 an, die durch Marker M173 unterschieden wird. R1 ist in Europa und Westasien sehr verbreitet. Es wird angenommen, dass seine Verbreitung mit der Wiederbesiedelung von Nord-Eurasien nach der letzten Eiszeit zusammenhängt. Seine Hauptuntergruppen sind R1a (SRY1532) und R1b (M343). Ein isolierter Stamm der Y-Chromosomen, die anscheinend zu Haplogruppe R1b1* (P25) gehören, wurde in hoher Konzentration unter der einheimischen Bevölkerung von Nordkamerun in West-Zentralafrika gefunden. Es wird angenommen, dass dies eine prähistorische Rückwanderung einer alten protoeurasischen Bevölkerung nach Afrika darstellt. Einige Forscher haben zudem über Y-Chromosomen der Haplogruppe T in geringer Konzentration in einigen dieser kamerunischen Bevölkerungen berichtet, die eine eurasische Ähnlichkeit aufweisen.[2]
Einige Y-Chromosomen, die ihrerseits mit den kamerunischen R1b1* Chromosomen eng verwandt zu sein scheinen, werden in hoher Konzentration unter der modernen Bevölkerung in Ägypten gefunden. Viele moderne Bevölkerungen des Nordkamerun sprechen tschadische Sprachen, die als ein alter Zweig der afro-asiatischen Sprachfamilie eingestuft werden. Die ausgestorbene Sprache des Now der alten Ägypter gehörte derselben Sprachfamilie an.
Personen, deren Y-Chromosomen alle Mutationen an den internen Nullpunkten des Y-DNA-Baums unter Einbeziehung von M207 (der Haplogruppe R definiert) aufweisen, aber weder die Mutation M173 (definiert Haplogruppe R1) noch die Mutation M124 (definiert Haplogruppe R2) besitzen, werden Haplogruppe R* zugerechnet. Einige Fälle von Haplogruppe R* sind in den Proben der australischen Aborigines gefunden worden. Haplogruppe R* wurde auch in 10,3 % (10/97) einer Probe von Burusho und in 6,8 % (3/44) einer Probe von Kalash in Nordpakistan entdeckt.[3]
Untergruppen
[Bearbeiten | Quelltext bearbeiten]Die Untergruppen der Haplogruppe R mit ihrer unterscheidenden Mutation, nach ihrer Gliederung, veröffentlicht 2010 von der International Society of Genetic Genealogy (ISOGG):[4]
- R
- R1 (M173/P241, M306/S1,P225, P231, P233, P234, P236, P238, P242, P245, P286, P294)
- R1a (L62/M513, L63/M511, L145/M449, L146/M420) Typisch für Bevölkerungen von Mittel- und Osteuropa, Zentralasien und Südasien, mit einer mittleren Verbreitung in Westeuropa, Südwestasien und Südsibirien
- R1a1 (L120/M516, L122/M448, M459, SRY1532.2/SRY10831.2)
- R1a1a (M17, M198, M417, M512, M514, M515)
- R1a1a1 (M56)
- R1a1a2 (M157)
- R1a1a3 (M64.2, M87, M204)
- R1a1a4 (P98)
- R1a1a5 (PK5)
- R1a1a6 (M434)
- R1a1a7 (M458)
- R1a1a7a (M334)
- R1a1a7b (L260)
- R1a1a7
- R1a1a
- R1a1
- R1a1a (M17, M198, M417, M512, M514, M515)
- R1a
- R1a1 (L120/M516, L122/M448, M459, SRY1532.2/SRY10831.2)
- R1b (M343) Typisch für Bevölkerungen in Westeuropa, mit einer mittleren Verbreitung außerhalb Eurasiens und in Teilen von Afrika
- R1b1 (P25, L278)
- R1b1a (V88), wird in Zusammenhang mit der Ausbreitung der Tschadischen Sprachen (inkl. Altägyptisch) gesehen.
- R1b1a1 (M18)
- R1b1a2 (V8)
- R1b1a3 (V35)
- R1b1a3a (V7)
- R1b1a3
- R1b1a4 (V69)
- R1b1a
- R1b1b (P297)
- R1b1b1 (M73)
- R1b1b2 (L265, M269, S3, S10, S13, S17)
- R1b1b2a (L23/S141, L49, L150)
- R1b1b2a1 (L51/S167)
- R1b1b2a1a (L11/S127, L52, L151, P310/S129, P311/S128)
- R1b1b2a1a1 (M405/S21/U106)
- R1b1b2a1a1a (M467/S29/U198)
- R1b1b2a1a1b (P107)
- R1b1b2a1a1c (DYS439(Null)/L1/S26)
- R1b1b2a1a1d (L48/S162)
- R1b1b2a1a1d1 (L47)
- R1b1b2a1a1d1a (L44)
- R1b1b2a1a1d1a1 (L45, L46, L164)
- R1b1b2a1a1d1a
- R1b1b2a1a1d1
- R1b1b2a1a1d1a (L44)
- R1b1b2a1a1d2 (L148)
- R1b1b2a1a1d
- R1b1b2a1a1d1 (L47)
- R1b1b2a1a1e (L257)
- R1b1b2a1a1
- R1b1b2a1a2 (P312/S116)
- R1b1b2a1a2a (M65)
- R1b1b2a1a2b (M153)
- R1b1b2a1a2c (M167/SRY2627)
- R1b1b2a1a2d (S28/U152)
- R1b1b2a1a2d1 (M126)
- R1b1b2a1a2d2 (M160)
- R1b1b2a1a2d3 (L2/S139)
- R1b1b2a1a2d3a (L20/S144)
- R1b1b2a1a2d3
- R1b1b2a1a2d4 (L4)
- R1b1b2a1a2d
- R1b1b2a1a2e (L165/S68)
- R1b1b2a1a2f (L21/S145)
- R1b1b2a1a2f1 (M37)
- R1b1b2a1a2f2 (M222/USP9Y+3636)
- R1b1b2a1a2f3 (P66)
- R1b1b2a1a2f4 (L226/S168)
- R1b1b2a1a2f5 (L193/S176)
- R1b1b2a1a2f
- R1b1b2a1a2
- R1b1b2a1a
- R1b1b2a1a1 (M405/S21/U106)
- R1b1b2a1
- R1b1b2a1a (L11/S127, L52, L151, P310/S129, P311/S128)
- R1b1b2a
- R1b1b2a1 (L51/S167)
- R1b1b2
- R1b1b2a (L23/S141, L49, L150)
- R1b1b
- R1b1c (M335)
- R1b1
- R1b1a (V88), wird in Zusammenhang mit der Ausbreitung der Tschadischen Sprachen (inkl. Altägyptisch) gesehen.
- R1b
- R1b1 (P25, L278)
- R1
- R1a (L62/M513, L63/M511, L145/M449, L146/M420) Typisch für Bevölkerungen von Mittel- und Osteuropa, Zentralasien und Südasien, mit einer mittleren Verbreitung in Westeuropa, Südwestasien und Südsibirien
- R2 (M124) Typisch für Bevölkerungen von Südasien, mit einer mittleren Verbreitung in Zentralasien und dem Kaukasus
Haplogruppe R1 macht die Mehrheit der Haplogruppe R in Form seiner Untergruppen, R1a und R1b, aus.[5][6][7]
R1a
[Bearbeiten | Quelltext bearbeiten]Die höchsten Konzentrationen von R1a (> 50 %) finden sich heute entlang der Eurasischen Steppe bei den oiratisch-sprachigen (ursprünglich turksprachigen) Khoton in der mongolischen Provinz von Uws (82,5 %), den Kirgisen (63,0 %), den Ishkashimi-Sprechern im Grenzbereich von Afghanistan, Pakistan und Tadschikistan (68 %), den Tadschiken von Khojant (64 %), den Sorben (63,39 %), den Schoren (58,8 %), den Polen (56,4 %), den Teleuten (55,3 %), den Ukrainern (54,0 %), den Süd-Altaiern (60,0 %) sowie in Ostindien unter den Brahmanen Westbengalens (72 %).[8][9][10][6][11][12][13][14]
In Deutschland sind 30 % aller Männer Merkmalsträger von R1a. Geschichtlich tritt eine R1a (M198+, Z93-) um 2500 v. Chr. in der bronzezeitlichen Nekropole Xiaohe (um 2000 v. Chr.) im nordchinesischen Xinjiang und bei den Tarim-Mumien (um 2900 v. Chr.) auf, die den geschichtlichen Tocharern entsprechen. Alle diese Funde hatten einheimische asiatische Mütter. Zur gleichen Zeit kam R1a ab etwa 2900 v. Chr. auch zahlreich in Schnurkeramiker-Gruppen von der russisch-weißrussischen Grenze bis nach Deutschland, zu den dänischen Inseln und Südschweden sowie in der späten Jamnaja-Kultur und der Aunjetitzer Kultur vor.[15]

R1a wurde von einzelnen Forschern in folgende geschichtliche Zusammenhänge gestellt:
- eine mögliche Neubesiedlung Eurasiens aus dem Refugium in der Ukraine Ende der letzten Eiszeit[16][6],
- die Expansion der Kurgankultur aus der ponto-kaspischen Steppe, die mit der Verbreitung der indoeuropäischen Sprachen einherging.[9][6]
R1b
[Bearbeiten | Quelltext bearbeiten]Die Haplogruppe R1b ist die wichtigste Haplogruppe in Westeuropa und im südlichen Ural. In Deutschland tragen 47 % der Männer R1b. Haplogruppe R1b entstand im südlichen Sibirien.[17]
Ursprung
[Bearbeiten | Quelltext bearbeiten]Die Y-Haplogruppe R1b ist eine Abzweigung von R1 (M173) und wird durch den Marker M343 unterschieden.[18] Der R1b-Stamm scheint einen viel höheren Grad an innerer Diversität als R1a zu haben, was nahelegt, dass die Mutation M343, die R1b von R1* trennt, beträchtlich früher aufgetreten sein muss als die Mutation SRY1532, die R1a von den übrigen R1-Ausprägungen unterscheidet. Das Genographic Project (geleitet von der National Geographic Society),[5] nimmt an, dass R1b auf der Iberischen Halbinsel entstanden ist, als diese für die damaligen Europäer ein Refugium während der Würmeiszeit bildete, und sich in der Folge von dort ausbreitete. Von dieser These wird inzwischen jedoch vermehrt Abstand genommen, da die Varianz (z. B. laut Barbara Arredi und Kollegen) in Eurasien wesentlich höher ist und nach Westeuropa hin kontinuierlich abnimmt, wobei zugleich die Menge der SNPs zunimmt. Dies deutet darauf hin, dass die iberische Population wesentlich jünger ist. Dabei bildet auch die (bis in die Jungsteinzeit zurückverfolgbare) baskische R1b-Population keine Ausnahme.
Inzwischen geht man vermehrt davon aus, dass die Westeuropäer auf keinen Fall älter als 10.000 Jahre sind und vor diesem Zeitpunkt entweder in Afrika, oder in Asien lebten. Von afrikanischen Wissenschaftlern wurde die These geäußert, dass sich die späteren Westeuropäer in der damals noch fruchtbaren Sahara aufhielten und durch das Austrocknen der Sahara über das Mittelmeer nach Südeuropa einwanderten. Dafür spricht die Subclade V88, die eindeutig mit der Ausbreitung der tschadischen Sprachen verbunden wird. Zumindest im Neuen Reich Ägyptens war R1b dort vertreten. Russische Wissenschaftler sind dagegen der Meinung, dass sich R1b vom Altai aus ausbreitete und seine Merkmalsträger ursprünglich eine Turksprache sprachen. Sie schließen R1b als Träger der indogermanischen Ursprache aus. Dieser These widerspricht jedoch die Existenz uralter R* und R1* im östlichen Iran, Pakistan, Afghanistan und Indien, wenn auch in kleiner Menge. Die Frage, wie R1b genau nach Europa kam, ist daher nach wie vor nicht eindeutig zu beantworten.
Heutige Verbreitung
[Bearbeiten | Quelltext bearbeiten]In Europa ist R1b (mit den Untergruppen R1b1 und R1b3, früher Hg1 und Eu18 genannt) die häufigste Y-Haplogruppe. Die Konzentration erreicht in Teilen des nordwestlichen Irland 98 %, in Nord- und Westengland, Spanien[6], Portugal und Irland[7] bis zu 90 % und im südöstlichen England und den Niederlanden[6] noch etwa 70 %. Darüber hinaus ist R1b in einigen Teilen Algeriens mit etwa 10 % vertreten.[19]
90 % der R2-Träger werden auf dem Indischen Subkontinent gefunden. R2 wurde ebenfalls im Kaukasus und in Zentralasien entdeckt.
R2 (M124) entstand vor ungefähr 25.000 Jahren im südlichen Zentralasien. Seine Träger wanderten nach Süden als Teil der zweiten[20] großen Auswanderungswelle nach Indien (die erste Einwanderung von Menschen auf den Subkontinent vor etwa 50.000 oder 60.000 Jahren hatte aus afrikanischen Migranten bestanden, die stets entlang der Küsten gewandert waren). Die fast ausschließliche Existenz von R2 in Indien könnte auch bedeuten, dass die Verzweigung von R in R1 und R2 in Mittelindien oder in Westindien geschah.
Siehe auch
[Bearbeiten | Quelltext bearbeiten]Literatur
[Bearbeiten | Quelltext bearbeiten]- Luigi Luca Cavalli-Sforza: The History and Geography of Human Genes. Princeton University Press, 1994. ISBN 0-691-08750-4.
- O. Semino et al.: The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective. In: Science. Volume 290, Issue 5494, 10. November 2000, S. 1155–1159, doi:10.1126/science.290.5494.1155.
- Wells et al.: The Eurasian Heartland: A continental perspective on Y-chromosome diversity. 2001 (Online [PDF; 2,3 MB; abgerufen am 27. Oktober 2021]).
- Passarino et al.: Different genetic components in the Norwegian population revealed by the analysis of mtDNA and Y chromosome polymorphisms. 2002 (nature.com).
- Behar et al.: Multiple Origins of Ashkenazi Levites: Y Chromosome Evidence for Both Near Eastern and European Ancestries. 2003 (uchicago.edu).
- Saha Anjana, Sharma Swarkar, Bhat Audesh, Pandit Awadesh, Bamezai Ramesh: Genetic affinity among five different population groups in India reflecting a Y-chromosome gene flow. In: J Hum Genet 50, 2005, S. 49–51, PMID 15611834
- Sanghamitra Sengupta et al.: Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists. In: American Journal of Human Genetics. 78, 2006, S. 202–221.
- C. Cinnioglu et al.: Excavating Y-chromosome haplotype strata in Anatolia. (PDF) In: Human Genetics. 114 (2), 2004, S. 127–148.
Weblinks
[Bearbeiten | Quelltext bearbeiten]- ISOGG tree of haplogroup R
- R branch of the haplotree
- 2005 Y-Chromosome Phylogenetic Tree
- World and European haplogroup prevalence maps (PDF; 386 kB)
- Spread of Haplogroup R1, from The Genographic Project, National Geographic (18. September 2010 disabled / offline)
- Spread of R1a1, from the Genographic Project, National Geographic (18. September 2010 disabled / offline)
- Spread of R1b, from the Genographic Project, National Geographic
- World Families Network zu R1b
- World Families Network zu R1a
- Worldfamilies.net Page on Haplogroups
{kind=link}
Einzelnachweise
[Bearbeiten | Quelltext bearbeiten]- ↑ Pille Hallast, Anastasia Agdzhoyan, Oleg Balanovsky, Yali Xue, Chris Tyler-Smith: A Southeast Asian origin for present-day non-African human Y chromosomes. In: Human Genetics. Band 140, Nr. 2, 1. Februar 2021, ISSN 1432-1203, S. 299–307, doi:10.1007/s00439-020-02204-9, PMID 32666166, PMC 7864842 (freier Volltext).
- ↑ C. A. Mulcare, M. E. Weale, A. L. Jones, B. Connell, D. Zeitlyn, A. Tarekegn, D. M. Swallow, N. Bradman, M. G. Thomas: The T allele of a single-nucleotide polymorphism 13.9 kb upstream of the lactase gene (LCT) (C-13.9kbT) does not predict or cause the lactase-persistence phenotype in Africans. In: American Journal of Human Genetics. Band 74, Nummer 6, Juni 2004, S. 1102–1110, doi:10.1086/421050, PMID 15106124, PMC 1182074 (freier Volltext).
- ↑ Sadaf Firasat, Shagufta Khaliq, Aisha Mohyuddin, Myrto Papaioannou, Chris Tyler-Smith, Peter A Underhill, Qasim Ayub: Y-chromosomal evidence for a limited Greek contribution to the Pathan population of Pakistan. In: European Journal of Human Genetics. (2007) 15, S. 121–126.
- ↑ isogg.org ISOGG Website
- ↑ a b Haplogroup R1 (M173). In: The Genographic Project. National Geographic Society, archiviert vom am 17. November 2006; abgerufen am 11. März 2008. Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.
- ↑ a b c d e f Semino et al. 2000.
- ↑ a b Rosser et al. 2000.
- ↑ Peričić et al.: High-Resolution Phylogenetic Analysis of Southeastern Europe Traces Major Episodes of Paternal Gene Flow Among Slavic Populations. In: Molecular Biology and Evolution, Band 22, Heft 10 (1. Oktober 2005), s. 1964–1975 (doi:10.1093/molbev/msi185).
- ↑ a b Wells et al. 2001.
- ↑ Behar et al. 2003.
- ↑ S. Sharma, E. Rai, S. Singh, P.R. Sharma, A.K. Bhat, K. Darvishi, A.J.S. Bhanwer, P.K. Tiwari, R.N.K. Bamezai: The Autochthonous Origin and a Tribal Link of Indian Brahmins: Evaluation Through Molecular Genetic Markers. In: The American Society of Human Genetics, 57th Annual Meeting, October 23–27, 2007, San Diego CA, S. 273 (1344/T).
- ↑ T. Katoh, B. Munkhbat, K. Tounai, S. Mano, H. Ando, G. Oyungerel, G. T. Chae, H. Han, G. J. Jia, K. Tokunaga, N. Munkhtuvshin, G. Tamiya, H. Inoko: Genetic features of Mongolian ethnic groups revealed by Y-chromosomal analysis. In: Gene. Band 346, Februar 2005, S. 63–70, doi:10.1016/j.gene.2004.10.023, PMID 15716011.
- ↑ Miroslava Derenko et al.: Contrasting patterns of Y-chromosome variation in South Siberian populations from Baikal and Altai-Sayan regions. (PDF; 441 kB) 2005
- ↑ V. N. Khar’kov: Gene pool differences between Northern and Southern Altaians inferred from the data on Y-chromosomal haplogroups. In: Genetika. 4, 3 (5) 2005, S. 675–687, PMID 17633562.
- ↑ Li 2010; Adler 2012; Brandt 2013; Haak 2015; Allentoft 2015.
- ↑ Passarino et al. 2002.
- ↑ Variations of R1b Ydna in Europe: Distribution and Origins. Архивировано из первоисточника 25 марта 2012.
- ↑ Note that in earlier literature the M269 marker, rather than M343, was used to define the R1b haplogroup. Then, for a time (from 2003 to 2005) what is now R1b1c was designated R1b3.
- ↑ Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample
- ↑ The Genographic Project. National Geographic Society, abgerufen am 13. März 2008.
| Evolutionsbaum Haplogruppen Y-chromosomale DNA (Y-DNA) | ||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adam des Y-Chromosoms | ||||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0’1'2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1’2'3’4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A1 | A2’3'4 | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2’3 | A4=BCDEF | |||||||||||||||||||||||||||||||||||||||||||||||||
| A2 | A3 | B | CT | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | |||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| G | IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| G1 | G2 | IJ | K | |||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | L | K(xLT) | T | ||||||||||||||||||||||||||||||||||||||||||||||
| | | | | | | ||||||||||||||||||||||||||||||||||||||||||||||||
| I1 | I2 | J1 | J2 | M | NO | P | S | |||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | | | |||||||||||||||||||||||||||||||||||||||||||||||||
| N | O | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1 | R2 | |||||||||||||||||||||||||||||||||||||||||||||||||
| | | ||||||||||||||||||||||||||||||||||||||||||||||||||
| R1a | R1b | |||||||||||||||||||||||||||||||||||||||||||||||||