Komagataeibacter
| Komagataeibacter | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Wildtyp von K. hansenii ATCC 23769, | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Komagataeibacter | ||||||||||||
| Yamada et al. 2013[2] |

Komagataeibacter ist eine Gattung von Essigsäurebakterien aus der Familie Acetobacteraceae,[4][5] einer Gruppe gramnegativer aerober Bakterien, die während der Gärung Essigsäure produzieren.[6] Sie wurde 2011/2012 von Yuzo Yamada et al. erstbeschrieben.[2][7] Die Typusart ist Komagataeibacter xylinus.[5]
Beschreibung[Bearbeiten | Quelltext bearbeiten]
Viele Komagataeibacter-Stämme (beispielsweise von der Spezies K. xylinus) sind eine Besonderheit unter den Essigsäurebakterien, da sie in der Lage sind Cellulose zu produzieren. Diese bakterielle Cellulose (BC), manchmal auch als (bakterielle) Nanocellulose (BNC)[1] bezeichnet, ist an der Bildung von Biofilmen beteiligt.[8] Sie ist zwar chemisch identisch mit pflanzlicher Cellulose, hat aber eine andere physikalische Struktur und damit andere physikalische Eigenschaften.[6] Sie ist ein widerstandsfähiges Material, das in der Medizin, der Geweberegeneration, bei Textilien, in der Elektronik und in der Biotechnologie im Allgemeinen eine Vielzahl von Verwendungszwecken und Anwendungen findet.[9]
Kombucha[Bearbeiten | Quelltext bearbeiten]


Eine weitere Anwendung ist die Herstellung von Kombucha durch die Fermentierung von gezuckertem Tee mit einer symbiotischen Kultur aus Bakterien und Hefen (Symbiotic Culture of Bacteria and Yeast, SCOBY);[9] diese Kombucha-Kulturen werden hier auch als mikrobielle Kombucha-Gemeinschaft (Kombucha Microbial Community, KMC) bezeichnet. In ihnen sind unter den Bakterien Komagataeibacter-Arten vorherrschend und produzieren dort Cellulose.[9] Die Masse aus mikrobiellen Kulturen und Cellulose wird analog zur Essigmutter beim Essig auch Kombuchamutter (Kombucha mother) genannt.
Etymologie[Bearbeiten | Quelltext bearbeiten]
Der Gattungsname Komagataeibacter ehrt den japanischen Mikrobiologen Kazuo Komagata (1928–2022),[10] Professor an der Universität Tokio, für seine Beiträge zur Systematik der Bakterien, insbesondere von Essigsäurebakterien.[5]
Artenliste[Bearbeiten | Quelltext bearbeiten]

Klade 1 (blau), Klade 2 (rot+grün), darin Klade 3 (grün) und Klade 4 (rot). Außengruppe: Acetobacter aceti (schwarz).
Synonyme des Gattungsnamens Komagataeibacter Yamada et al. 2013 sind nach der List of Prokaryotic names with Standing in Nomenclature (LPSN):[5][4]
- „Komagatabacter“ Yamada et al. 2012 (Schreibvariante)[7]
- Novacetimonas Brandão et al. 2022

Artenliste nach der LPSN[5] – mit einigen Stämmen (wo nicht anders angegeben) nach der Taxonomie des National Center for Biotechnology Information (NCBI),[11] Stand 22. Oktober 2023:
- Komagataeibacter cocois Liu et al. 2018[12]
- Referenzstamm: WE7 alias CGMCC 1.15338 oder JCM 31140[12]
- Komagataeibacter diospyri Naloka et al. 2020[13]
- Komagataeibacter europaeus (Sievers et al. 1992) Yamada et al. 2013[14]
- Komagataeibacter hansenii (Gosselé et al. 1983) Yamada et al. 2013
- Komagataeibacter intermedius (Boesch et al. 1998) Yamada et al. 2013[19]
- Komagataeibacter kakiaceti (Iino et al. 2012) Yamada 2014[20]
- weitere Stamm: NBRC 3288
- Komagataeibacter kombuchae (Dutta & Gachhui 2007) Yamada et al. 2013[A. 1]
- Referenzstamm: RG3 alias LMG 23726 oder MTCC 6913
- Komagataeibacter maltaceti (Slapšak et al. 2013) Yamada 2014[20]
- Komagataeibacter medellinensis (Castro et al. 2013) Yamada 2014[20]
- Komagataeibacter melaceti Marič et al. 2020
- Referenzstamm: AV382 alias ZIM:B1054, CCM:8958 oder BCCM/LMG:31303
- Komagataeibacter melomenusus Marič et al. 2020
- Referenzstamm: AV436 alias ZIM:B1056, BCCM/LMG:31304 oder CCM:8959
- Komagataeibacter nataicola (Lisdiyanti et al. 2006) Yamada et al. 2013[15]
- Referenzstamm: JCM:25120 alias BCCM/LMG:1536 oder NRIC:0616
- weiterer Stamm: RZS01[15]
- Komagataeibacter oboediens (Sokollek et al. 1998) Yamada et al. 2013[9]
- Komagataeibacter pomaceti Škraban et al. 2019[21]
- Referenzstamm: BCC 36444 alias JCM:25121, BCCM/LMG:1582 oder NRIC:0614
- Komagataeibacter rhaeticus (Dellaglio et al. 2005) Yamada et al. 2013
- Komagataeibacter saccharivorans (Lisdiyanti et al. 2006) Yamada et al. 2013
- Referenzstamm: BCC 36444 alias JCM:25121, BCCM/LMG:1582 oder NRIC:0614
- weiterer Stamm: CV1[9]
- Komagataeibacter sucrofermentans (Toyosaki et al. 1996) Yamada et al. 2013
- Referenzstamm: BPR 2001 alias DSM:15973,[3] JCM:9730 oder BCCM/LMG:18788
- Komagataeibacter swingsii (Dellaglio et al. 2005) Yamada et al. 2013
- Referenzstamm: DST GL01 alias DST GL_01, DSM:16373, JCM:17123 oder BCCM/LMG:22125
- Komagataeibacter xylinus (Brown 1886) Yamada et al. 2013 (Typusart)[A. 2]
- Komagataeibacter sp. K2A8[3][27]
Komagataeibacter xylinus[Bearbeiten | Quelltext bearbeiten]
Komagataeibacter xylinus ist eine Bakterienart, deren Bedeutung vor allem darin liegt, dass sie Cellulose (Bakteriencellulose oder Nanocellulose) produzieren kann.[A. 2]
Der optimale pH-Wert dieser Bakterien liegt bei 5,4 bis 6,3 und die optimale Temperatur für die Kultivierung bei 25 bis 30 °C. Außerhalb dieses pH- und Temperaturbereichs verlangsamt sich das Bakterienwachstum, starke Abweichung kann sogar zum Absterben der Bakterien führen.[28]
Forschungsgeschichte und Systematik[Bearbeiten | Quelltext bearbeiten]
Die Art (Spezies) wurde erstmals 1886 von Adrian John Brown beschrieben, der das Bakterium bei Untersuchungen zur Fermentation identifizierte. Brown gab der Art den Namen Bacterium xylinum. (Bacterium war ursprünglich einmal eine Gattungsbezeichnung). Seitdem ist die Art unter verschiedenen anderen Namen bekannt, vor allem Acetobacter xylinum und Gluconacetobacter xylinus.[8] Ihren heutigen Namen erhielt sie 2012 mit der Etablierung der neuen Gattung Komagataeibacter[8][7][2] als Typusart der Gattung.[5]
Genom und Stoffwechsel[Bearbeiten | Quelltext bearbeiten]
Das Genom des Cellulose-defizienten[A. 3] Stammes K. xylinus NBRC 3288 wurde 2011 sequenziert,[26] gefolgt von den Genomen Zellulose-produzierender Stämme in den Jahren 2014 (Stamm E25)[24] und 2018 (Stamm CGMCC 2955).[25] Der erste dieser beiden Zellulose-produzierende Stämme (E25) hat ein Genom, das aus einem Chromosom mit 3,4 Megabasenpaaren und fünf Plasmiden besteht, von denen eines ein „Megaplasmid“ mit etwa 330 Kilobasenpaaren ist.[24] Die Schlüsselgene für die Celluloseproduktion befinden sich im Vier-Gen-Operon bcsABCD, das für die vier Untereinheiten des Enzyms Cellulose-Synthase kodiert. Alle vier Gene sind für eine effiziente Celluloseproduktion in vivo erforderlich, wobei BcsA und BscB in vitro ausreichend sind. Mehrere andere Gene im Genom von K. xylinus sind ebenfalls an der Celluloseproduktion und -regulierung beteiligt, darunter ein Cellulase-Enzym.[8]
Bedeutung und Anwendungen[Bearbeiten | Quelltext bearbeiten]

.jpg)

K. xylinus wurde lange Zeit als Modellorganismus für die Untersuchung der Celluloseproduktion in Pflanzen verwendet. Die Spezies dient ebenfalls zur Untersuchung der bakteriellen Biofilmproduktion, der Zell-Zell-Kommunikation und anderer interessanter Themen.[8] Die Produktion von bakterieller Cellulose (BC) für industrielle Zwecke ist Gegenstand umfangreicher Forschungsarbeiten, ist jedoch (noch) begrenzt bzgl. Produktivität und Skalierbarkeit begrenzt (Stand 2018).[6][25]
Eine weitere potenzielle Anwendung für K. xylinus ist die Herstellung von Papier: die Spezies könnte zur Herstellung von starkem Papier ohne Holz verwendet werden. Das in einem energieeffizienten und umweltfreundlichen Verfahren hergestellte Material könnte als alternatives Verpackungsmaterial zum Frischhalten von Lebensmitteln verwendet werden oder für Trinkpäckchen, Lautsprechermembranen, zur Wundversorgung etc. Im Jahr 2007 gewann dieser Vorschlag von Mareike Frensemeier für eine mögliche Anwendung mit dem Titel Bacs den dritten Platz beim Bayerischen MaterialScience VisionWorks Award.[29]
K. xylinus (früher Acetobacter xylinus) ist der wichtigste Mikroorganismus in der Kombucha-Kultur[30] und wird auf den Philippinen traditionell auch für die Herstellung der geleeartigen Desserts Nata de Piña[31] und Nata de Coco verwendet, die aus Ananassaft respektive Kokosnusswasser hergestellt werden. Die erstere Variante wird bereits seit dem 18. Jahrhundert hergestellt.[32][33]
Komagataeibacter hansenii[Bearbeiten | Quelltext bearbeiten]


Komagataeibacter hansenii ist eine weitere Spezies der Essigsäurebakterien, die als Modellorganismus für die Biosynthese von bakterieller Cellulose dient,[34][35] der Stamm ATCC 23769 wurde vollständig sequenziert.[15] Der Stamm CGMCC 3917 von K. hansenii (alias Gluconacetobacter hansenii) kann bakterielle Cellulose produzieren, wobei ihm lediglich Abfälle an Bierhefe als Nahrungsquelle dienen.[16]
In vielen Ländern (insbesondere auch in China) ist die Bierherstellung ein wichtiger Wirtschaftszweig. Dort können entsprechend große Mengen an Bierhefeabfällen (waste beer yeasts, WBY) als Nebenprodukt der Brauereiindustrie anfallen. Diese werden, soweit sie nicht an Vieh und Geflügel verfüttert werden, weggeworfen, was nicht nur zu einer Reihe von Umweltproblemen führt, sondern auch eine enorme Verschwendung von Ressourcen darstellt. Cellulose ist ein wichtiger Rohstoff mit vielerlei Anwendungen. Dabei zeigt bakterielle gegenüber pflanzlicher Cellulose viele ungewöhnliche physikalisch-chemische und mechanische Besonderheiten. Dazu gehören u. a. eine höhere Reinheit, eine höhere Wasseraufnahme- und -haltekapazität; ein höherer Polymerisationsgrad geht einher mit einer höheren Zugfestigkeit. Die mikrobiologische Produktion von dieser besonderen Form der Cellulose könnte daher eine neue besonders nützliche Lösung des Entsorgungsproblems der überschüssigen Bierhefeabfälle sein.[16]
Komagataeibacter nataicola[Bearbeiten | Quelltext bearbeiten]
Komagataeibacter nataicola ist eine Spezies, die reichlich bakterielle Cellulose (BC) produzieren kann und hohe Konzentrationen von Essigsäure toleriert. Das Genom des Cellulose-produzierenden Stammes RZS01 wurde 2017 von Zhang et al. vollständig sequenziert. Es besteht aus einem Chromosom von 3.485.191 bp (Basenpaaren), dazu kommen 6 Plasmide mit 25.766 bp bis 102.282 bp. Es gibt im Genom vorhergesagt 3.609 Offene Leserahmen (open reading frames, ORFs), die für 3.514 Proteine kodieren und drei Cellulose-Synthase-Operons tragen.[15]
Die Spezies findet in Neuseeland (aufgrund ihrer Säureverträglichkeit) Anwendung zur Herstellung von Tamarillo-Essig (Tamarillo Vinegar).[36][37][38]
Komagataeibacter oboediens[Bearbeiten | Quelltext bearbeiten]

RhodPhage: „Rhodovulum-Phage RS1“
RuegPhage: „Ruegeria phage DSS3-P1“
Die Spezies Komagataeibacter oboediens scheint unter Bedingungen, wie sie auf dem Mars herrschen, überleben zu können.
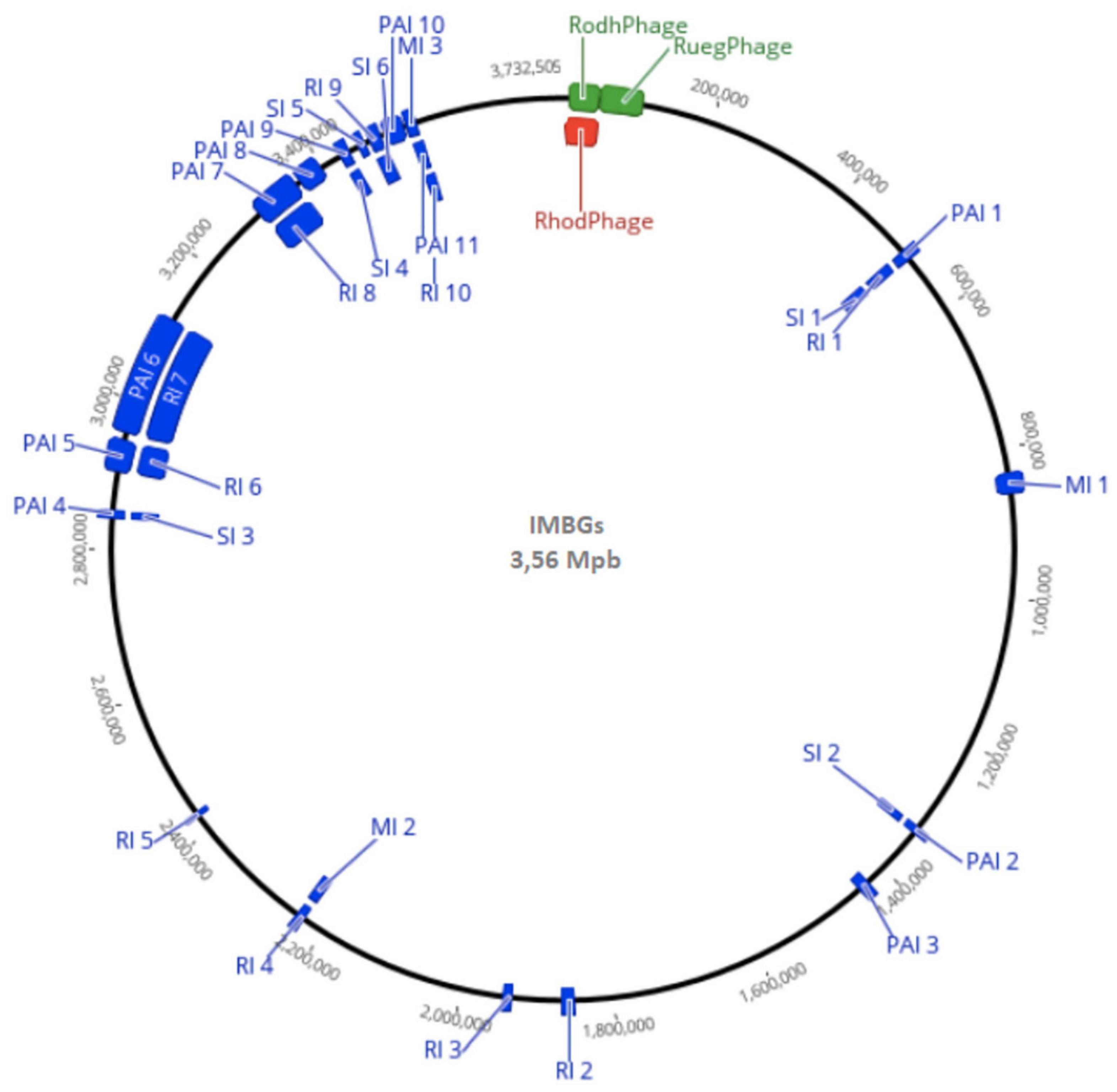
In einer 2022 veröffentlichten Studie hatten Daniel Santana de Carvalho et al. untersucht, ob Kombucha-Kulturen unter marsähnlichen Bedingungen überleben können. Dazu wurde der Cellulose-produzierenden Stamm K. oboediens IMBG180 auf der Internationalen Raumstation ISS marsähnlichen Bedingungen ausgesetzt, insbesondere einer simulierten Marsatmosphäre (Biology and Mars Experiment, BIOMEX). Obwohl die simulierte Marsatmosphäre die mikrobielle Ökologie der Kombucha-Kulturen zerstörte, überlebten die Bakterien von K. oboediens die Prozedur.[9] Dies bestätigte im Jahr 2021 gewonnene Ergebnisse am Stamm IMabG 314.[22] Um zu erfahren, welche Veränderungen am Genom die unter den extremen Bedingungen ausgesetzten Bakterien erfuhren, wurde das Genom des ursprünglichen Stammes mit der aus der Exposition hervorgegangene und mit IMBG185 bezeichneten Variante verglichen. Tatsächlich zeigte das Genom von K. oboediens nach Reaktivierung auf der Erde nur geringe Veränderungen. Es scheint, dass die von den Bakterien produzierte Cellulose ihr Überleben unter den extraterrestrischen Bedingungen ermöglicht hat. Membranen oder Filme auf Basis bakterieller Cellulose (BC) könnten daher ein geeignetes Biomaterial für den Schutz von Leben in extraterrestrischen Siedlungen sein. Darüber hinaus ist dies der erste Beleg dafür, dass bakterielle Zellulose ein Biomarker für extraterrestrisches Leben sein könnte.[9]
Prophagen[Bearbeiten | Quelltext bearbeiten]
Die Genomanalysen der beiden IMBG-Varianten zeigten auch 96 Kandidaten für Prophagen, darunter drei (vielleicht vier) aktive. Diese zeigten Ähnlichkeiten mit folgenden Bakteriophagen:[9][A. 4]
- „Rhodovulum-Phage RS1“ (Caudoviricetes, vom Morphotyp der Siphoviren)[39][40]
- „Ruegeria-Phage DSS3-P1“ (Casjensviridae, vom Morphotyp der Siphoviren)[41]
- ein nicht näher bezeichneter Streptomyces-Phage
Komagataeibacter europaeus[Bearbeiten | Quelltext bearbeiten]
Die Art Komagataeibacter europaeus ist eine der am Essigsäuregärungsprozess (acetic acid fermentation, AAF) von traditionellem chinesischem Getreideessig (cereal vinegar, →Reisessig) minder stark vertretene Spezies, deren Wirkung dennoch für das Gesamtergebnis von entscheidender Bedeutung ist.[14]
Diese Fermentation wird erzielt durch is komplexes und vielfältige Ökosystem, an dem mehrere Spezies von Mikroorganismen beteiligt sind, darunter einige mehr und einige weniger verbreitete Arten. Die Rolle der letzteren bei der Essiggärung war jedoch lange Zeit nicht sehr gut erforscht. In einer 2021 veröffentlichten Studie untersuchten Ming-Ye Peng et al. solche Mischkulturen mit weniger stark vorhandenen bakteriellen Untergemeinschaften anhand der traditionellen aromatischen Essiggärung der chinesischen Provinz Zhenjiang als Modellsystem. Die an dieser Fermentation weniger stark vertretenen Vertreter – wie K. europaeus – reagierten empfindlich auf Veränderungen der Umwelt, insbesondere des Zuckergehalts und der Gesamtsäurekonzentration, spielten jedoch insgesamt eine grundlegende Rolle für die Stabilität des gesamten mikrobiellen Netzwerks. Dies zeigte ihre Bedeutung für die metabolische Funktion und die Widerstandsfähigkeit der untersuchten mikrobiellen Gemeinschaft hindeutet, weshalb diesen eine wichtige ökologische Rolle in der Produktion traditioneller fermentierter Lebensmitteln zukommt. In der Studie wurde untersucht, wie der untersuchte Fermentierungsprozess durch Bioaugmentation von K. europaeus JNP1 die Fermentierung optimiert werden kann. Die Studie unterstreicht die Bedeutung auch weniger stark vertretener Arten als bakterielle Keimbank (seed bank), um die biologische Vielfalt der Essigmikrobiota zu erhalten.[14]
Siehe auch[Bearbeiten | Quelltext bearbeiten]
Weblinks[Bearbeiten | Quelltext bearbeiten]
- Guruprakash Subbiahdoss, Sarah Osmen, Erik Reimhult: Cellulosic biofilm formation of Komagataeibacter in kombucha at oil-water interfaces. In: Biofilm, Band 4, 26. Februar 2022, S. 100071; doi:10.1016/j.bioflm.2022.100071, ePub Dezember 2022 (englisch).
- Claudia Krapp: Wie Bakterien veganes Leder produzieren und färben. Auf: wissenschaft.de vom 5. April 2024.
Anmerkungen[Bearbeiten | Quelltext bearbeiten]
- ↑ a b Der Name Komagataeibacter kombuchae (mit Referezstamm RG3) wird nach der LPSN von vielen Autoren – wie auch in der NCBI-Taxonomie – als ein Synonym von K. hansenii gesehen (und RG3 als einer er Stämme dieser Art), nicht jedoch in der LPSN selbst.
- ↑ a b In der GTDB ist die Spezies Komagataeibacter xylinus dreifach aufgeteilt: Von der eigentlichen Art K. xylinus mit Referenzstamm NBRC 15237 und den beiden weiteren Stämmen LMG 1515 und DSM 2325[9] abgetrennt sind die Spezies mit den provisorischen Bezeichnungen Komagataeibacter xylinus_C (Referenzstamm K2G30[3]) und K. xylinus_D (Referenzstamm CGMCC 17276).[23]
- ↑ Cellulose-defizient = kann keine Cellulose produzieren
- ↑ Die beiden identifizierten Phagenspezies sind derzeit (Oktober 2023) noch nicht vom International Committee on Taxonomy of Viruses (ICTV) bestätigt, zum Streptomyces-Phagen liegen keine Angaben vor.
Einzelnachweise[Bearbeiten | Quelltext bearbeiten]
- ↑ a b c d Paulina Jacek, Małgorzata Ryngajłło, Stanisław Bielecki: Structural changes of bacterial nanocellulose pellicles induced by genetic modification of Komagataeibacter hansenii ATCC 23769. In: Applied microbial and cell physiology, Band 103, 29. Aril 2019, S. 5339–5353; doi:10.1007/s00253-019-09846-4 (englisch).
- ↑ a b c Yuzo Yamada, Pattaraporn Yukphan, Huong Thi Lan Vu, Yuki Muramatsu, Duangjai Ochaikul, Somboon Tanasupawat, Yasuyoshi Nakagawa: Description of Komagataeibacter gen. nov., with proposals of new combinations (Acetobacteraceae). In: The Journal of General and Applied Microbiology, Band 58, Nr. 5, November 2012, S. 397–404, doi:10.2323/jgam.58.397, PMID 23149685, ResearchGate (englisch).
- ↑ a b c d e Marcello Brugnoli, Francesco Robotti, Salvatore La China, Kavitha Anguluri, Hossein Haghighi, Simone Bottan, Aldo Ferrari, Maria Gullo: Assessing effectiveness of Komagataeibacter strains for producing surface-microstructured cellulose via guided assembly-based biolithography. In: Nature: Scientific Reports, Band 11, Nr. 19311, 29. September 2021; doi:10.1038/s41598-021-98705-2 (englisch).
- ↑ a b Aidan C. Parte: LPSN – List of Prokaryotic names with Standing in Nomenclature (bacterio.net), 20 years on. In: International Journal of Systematic and Evolutionary Microbiology. 68. Jahrgang, Nr. 6, 1. Juni 2018, ISSN 1466-5026, S. 1825–1829, doi:10.1099/ijsem.0.002786, PMID 29724269 (englisch).
- ↑ a b c d e f List of Prokaryotic names with Standing in Nomenclature (LPSN): Genus Komagataeibacter Yamada et al. 2013.
- ↑ a b c Dieter Klemm, Friederike Kramer, Sebastian Moritz, Tom Lindström, Mikael Ankerfors, Derek Gray, Annie Dorris: Nanocelluloses: A New Family of Nature-Based Materials. In: Angewandte Chemie International Edition. 50. Jahrgang, Nr. 24, 6. Juni 2011, S. 5438–5466, doi:10.1002/anie.201001273, PMID 21598362 (englisch).
- ↑ a b c Yuzo Yamada, Pattaraporn Yukphan, Huong Thi Lan Vu, Yuki Muramatsu, Duangjai Ochaikul, Yasuyoshi Nakagawa: Subdivision of the genus Gluconacetobacter Yamada, Hoshino and Ishikawa 1998: the proposal of Komagatabacter gen. nov., for strains accommodated to the Gluconacetobacter xylinus group in the α-Proteobacteria. In: Annals of Microbiology, Band 62, Nr. 2, 12. Juni 20112, ISSN 1869-2044, S. 849–859, doi:10.1007/s13213-011-0288-4 (englisch).
- ↑ a b c d e Ute Römling, Michael Y. Galperin: Bacterial cellulose biosynthesis: diversity of operons, subunits, products, and functions. In: Trends in Microbiology. 23. Jahrgang, Nr. 9, September 2015, S. 545–557, doi:10.1016/j.tim.2015.05.005, PMID 26077867, PMC 4676712 (freier Volltext) – (englisch).
- ↑ a b c d e f g h i j k l m n
Daniel Santana de Carvalho, Ana Paula Trovatti Uetanabaro, Rodrigo Bentes Kato, Flávia Figueira Aburjaile, Arun Kumar Jaiswal, Rodrigo Profeta, Rodrigo Dias De Oliveira Carvalho, Sandeep Tiwar, Anne Cybelle Pinto Gomide, Eduardo Almeida Costa, Olga Kukharenko, Iryna Orlovska, Olga Podolich, Oleg Reva6 Pablo Ivan P. Ramos, Vasco Ariston De Carvalho Azevedo, Bertram Brenig, Bruno Silva Andrade, Jean-Pierre P. de Vera, Natalia O. Kozyrovska, Debmalya Barh, Aristóteles Góes-Neto: The Space-Exposed Kombucha Microbial Community Member Komagataeibacter oboediens Showed Only Minor Changes in Its Genome After Reactivation on Earth. In: Frontiers in Microbiology, Band 13, Sec. Evolutionary and Genomic Microbiology, 11. März 2022; doi:10.3389/fmicb.2022.782175 (englisch). Siehe insbes. Fig. 4 (Prophagen). Dazu:
- Scientists Discover That Cellulose-Producing Bacteria Can Survive on Mars. Auf: SciTechDaily vom 9. August 2022. Quelle: Universität Göttingen.
- ↑ Mitsuo Sakamoto, Ken-ichiro Suzuki: In memoriam – Kazuo Komagata (1928–2022). In: International Journal of Systematic and Evolutionary Microbiology, Band 73, 005791, 2023; doi:10.1099/ijsem.0.005791, PDF (Nachruf, englisch).
- ↑ NCBI Taxonomy Browser: Komagataeibacter, details: Komagataeibacter Yamada et al. 2013 (genus).
- ↑ a b Long-xiang Liu, Si-xin Liu, Yan-mei Wang, Ji-cai Bi, Hua-mei Chen, Jian Deng, Cui Zhang, Qi-song Hu, Cong-fa Li: Komagataeibacter cocois sp. nov., a novel cellulose-producing strain isolated from coconut milk In: International Journal of Systematic and Evolutionary Microbiology., Band 68, Nr. 10, 22. August 2018, ISSN 1466-5026, S. 3125–3131, doi:10.1099/ijsem.0.002947, PMID 30132753 (englisch).
- ↑ a b c Kallayanee Naloka, Pattaraporn Yukphan, Minenosuke Matsutani, Kazunobu Matsushita, Gunjana Theeragool: Komagataeibacter diospyri sp. nov., a novel species of thermotolerant bacterial nanocellulose-producing bacterium. In: International Journal of Systematic and Evolutionarydddd Microbiology, Band 70, Nr. 1, 3. Februar 2020, ISSN 1466-5026, S. 251–258; doi:10.1099/ijsem.0.003745, PMID 31622229 (englisch).
- ↑ a b c d Ming-Ye Peng, Xiao-Juan Zhang, Ting Huang, Xiao-Zhong Zhong, Li-Juan Chai, Zhen-Ming Lu, Jin-Song Shi, Zheng-Hong Xu: Komagataeibacter europaeus improves community stability and function in solid-state cereal vinegar fermentation ecosystem: Non-abundant species plays important role. In: Food Research International, Band 150, Teil B, Dezember 2021, S. 110815; doi:10.1016/j.foodres.2021.110815 (englisch).
- ↑ a b c d e f g h i j k Heng Zhang, Xuran Xu, Xiao Chen, Fanshu Yuan, Bianjing Sun, Yunhua Xu, Jiazhi Yang, Dongping Sun: Complete genome sequence of the cellulose-producing strain Komagataeibacter nataicola RZS01, In: Nature: Scientific Reports, Band 7, Nr. 4431, 30. Juni 2017; doi:10.1038/s41598-017-04589-6 (englisch).
- ↑ a b c Dehui Lin, Patricia Lopez-Sanchez, Rui Li, Zhixi Li: Production of bacterial cellulose by Gluconacetobacter hansenii CGMCC 3917 using only waste beer yeast as nutrient source. In: Bioresource Technology, Band 151, 2014, S. 113–119; doi:10.1016/j.biortech.2013.10.052, PMID 24212131 (englisch).
- ↑ a b Małgorzata Ryngajłło, Marzena Jędrzejczak-Krzepkowska, Katarzyna Kubiak, Karolina Ludwicka & Stanisław Bielecki: Towards control of cellulose biosynthesis by Komagataeibacter using systems-level and strain engineering strategies: current progress and perspectives. In: Applied Microbiology and Biotechnology, Band 104, S. 6565–6585, 11. Juni 2020; doi:10.1007/s00253-020-10671-3 (englisch).
- ↑ Martin Bimmer, Wolfgang Liebl, Armin Ehrenreich: Bakterielle Cellulose — ein Netzwerk gestaltet von drei Cellulosesynthasen. In: BIOspektrum, Band 29, Biotechnologie: Cellulosesynthese, 13. April 2023, S. 218–220; doi:10.1007/s12268-023-1908-9, PDF (deutsch).
- ↑ a b Pietro Cannazza, Antti J. Rissanen, Essi Sarlin, Dieval Guizelini, Carlotta Minardi, Pauli Losoi, Francesco Molinari, Diego Romano, Rahul Mangayil: Characterization, genome analysis and genetic tractability studies of a new nanocellulose producing Komagataeibacter intermedius isolate. IN: Nature: Scientific Reports, Band 12, Nr. 20520, 28. November 2022; doi:10.1038/s41598-022-24735-z (englisch).
- ↑ a b c d e f Yuzo Yamada: Transfer of Gluconacetobacter kakiaceti, Gluconacetobacter medellinensis and Gluconacetobacter maltaceti to the genus Komagataeibacter as Komagataeibacter kakiaceti comb. nov., Komagataeibacter medellinensis comb. nov. and Komagataeibacter maltaceti comb. nov. In: International Journal of Systematic and Evolutionary Microbiology, Band 64, Nr. 5, 1. Mai 2014, ISSN 1466-5026, S. 1670–1672, doi:10.1099/ijs.0.054494-0, PMID 24523443 (englisch).
- ↑ a b Jure Škraban, Ilse Cleenwerck, Peter Vandamme, Lijana Fanedl, Janja Trček: Genome sequences and description of novel exopolysaccharides producing species Komagataeibacter pomaceti sp. nov. and reclassification of Komagataeibacter kombuchae (Dutta and Gachhui 2007) Yamada et al., 2013 as a later heterotypic synonym of Komagataeibacter hansenii (Gosselé et al. 1983) Yamada et al., 2013. In: Systematic and Applied Microbiology, Band 41, Nr. 6, 1. November 2018, ISSN 0723-2020, S. 581–592; doi:10.1016/j.syapm.2018.08.006, PMID 30177404 (englisch).
- ↑ a b Iryna Orlovska, Olga Podolich, Olga Kukharenko, Iryna Zaets, Oleg Reva, Ludmila Khirunenko, Danica Zmejkoski, Sergiy Rogalsky, Debmalya Barh, Sandeep Tiwari, Ranjith Kumavath, Aristóteles Góes-Neto, Vasco Azevedo, Bertram Brenig, Preetam Ghosh, Jean-Pierre de Vera, Natalia Kozyrovska: Bacterial Cellulose Retains Robustness but Its Synthesis Declines After Exposure to a Mars-like Environment Simulated Outside the International Space Station. In: Astrobiology. Band 21, Nr. 6, Juni 2021, S. 706–717; doi:10.1089/ast.2020.2332, Epub 26. Februar 2021 (englisch). Dazu:
- ↑ a b GTDB: Komagataeibacter (gen?).
- ↑ a b c Katarzyna Kubiak, Marta Kurzawa, Marzena Jędrzejczak-Krzepkowska, Karolina Ludwicka, Mariusz Krawczyk, Andrzej Migdalski, Magdalena M. Kacprzak, Damian Loska, Alina Krystynowicz, Stanisław Bielecki: Complete genome sequence of Gluconacetobacter xylinus E25 strain—Valuable and effective producer of bacterial nanocellulose. In: Journal of Biotechnology, Band 176, April 2014, S. 18–19, doi:10.1016/j.jbiotec.2014.02.006, PMID 24556328 (englisch).
- ↑ a b c Miao Liu, Lingpu Liu, Shiru Jia, Siqi Li, Yang Zou, Cheng Zhong: Complete genome analysis of Gluconacetobacter xylinus CGMCC 2955 for elucidating bacterial cellulose biosynthesis and metabolic regulation. In: Scientific Reports, Band 8, Nr. 1, 19. April 2018, ISSN 2045-2322, S. 6266, bibcode:2018NatSR...8.6266L; doi:10.1038/s41598-018-24559-w, PMID 29674724, PMC 5908849 (freier Volltext) (englisch).
- ↑ a b Hidetaka Ogino, Yoshinao Azuma, Akira Hosoyama, Hidekazu Nakazawa, Minenosuke Matsutani, Akihiro Hasegawa, Ken-ichiro Otsuyama, Kazunobu Matsushita, Nobuyuki Fujita, Mutsunori Shirai: Complete Genome Sequence of NBRC 3288, a Unique Cellulose-Nonproducing Strain of Gluconacetobacter xylinus Isolated from Vinegar. In: Journal of Bacteriology, Band 193, Nr. 24, 28. November 2011, S. 6997–6998; doi:10.1128/JB.06158-11, PMID 22123756, PMC 3232855 (freier Volltext) (englisch).
- ↑ NCBI Nzucleotide: Komagataeibacter xylinus strain K2A8 16S ribosomal RNA gene, partial sequence.
- ↑ Bernd H. A. Rehm: Microbial Production of Biopolymers and Polymer Precursors: Applications and Perspectives. Caister Academic, Januar 2009, 294 Seiten, ISBN 978-1-904455-36-3, SemanticScholar (englisch).
- ↑ VisionWorks Award CarsoPacks 2020. Bacs. Hochschule für Künste Bremen. Award Recipient Mareike Frensemeier. Memento im Webarchiv vom 20. September 2017.
- ↑ I. Jankovic, M. Stojanovic: Microbial and chemical composition, growth, therapeutical and antimicrobial characteristics of tea fungus. In: Mikrobiologija (Yugoslavia), 31. Oktober 1994; SemanticScholar (englisch).
- ↑ Nata de piña (cuisine philippine). Lexique culinaire, auf: gastronomiac.com (französisch).
- ↑ Agus Tri Sutanto: Pineapple Liquid Waste as Nata De Pina Raw Material. In: Makara, Teknologi. 16. Jahrgang, Nr. 1, Mai 2012, S. 63–67, doi:10.7454/mst.v16i1.1286 (englisch). ResearchGate, Academia.
- ↑ Benito S. Vergara, Panna Melizah H. Idowu, Julia H. Sumangil;Wilford Jan C. Almoro (Illustrations): Nata de Coco: A Filipino Delicacy. National Academy of Sciences and Technology, Philippines, 1999, ISBN 971-8538-61-5 (englisch, gov.ph [PDF]).
- ↑ Peter Ross, Raphael Mayer, Moshe Benziman: Cellulose biosynthesis and function in bacteria. In: ASM Journals: Microbiological Reviews, Band 55, Nr. 1, 1. März 1991, S. 35–58; doi:10.1128/mr.55.1.35-58.1991, PMID 2030672, PMC 372800 (freier Volltext), ResearchGate, Epub April 1991 (englisch).
- ↑ Ute Römling: Molecular biology of cellulose production in bacteria. In: Research in microbiology, Band 153, Nr. 4, Mai 2002, S. 205–212; doi:10.1016/S0923-2508(02)01316-5, EropePMC, Epub 29. März 2002 (englisch).
- ↑ GBIF: Komagataeibacter nataicola (Lisdiyanti et al., 2006) Yamada et al., 2013 – Occurrenc. 24. Mai 2012.
- ↑ Tamarillo Vinegar. Auf: Artisan Vinegar.
- ↑ Tamarillo recipes. Auf: Tamco, Neuseeland.
- ↑ Mara E. Heinrichs, Benedikt Heyerhoff, Berin S. Arslan-Gatz, Michael Seidel, Jutta Niggemann, Bert Engelen: Deciphering the Virus Signal Within the Marine Dissolved Organic Matter Pool. In: Frontiers in Microbiology, Band 13, Sec. Aquatic Microbiology, 27. Mai 2022; doi:10.3389/fmicb.2022.863686 (englisch).
- ↑ NCBI Taxonomy Browser: ncbi.nlm.nih.gov Rhodovulum phage RS1 (species).
- ↑ NCBI Taxonomy Browser: Ruegeria phage DSS3-P1 (species), synonm: Silicibacter phage DSS3-P1, Roseophage DSS3-P1.
{kind=link}